Využití chelatační afinitní chromatografie (IMAC) při studiu a
purifikaci biomolekul
Oldřich Janiczek,
Přírodovědecká fakulta MU, Brno
Obrovský rozvoj na
poli proteinové chemie v posledních padesáti letech byl podmíněn vývojem velmi efektivních
technik proteinové separace a analýzy. Převážná většina z nich je založena na rozdílné
hmotnosti či náboji separovaných biomolekul. V posledních
dvaceti letech však nabývá na významu skupina metod založených na specifické interakci
mezi biomolekulou a ligandem.
V případě, že je ligand imobilizován na vhodné stacionární
fázi a provádíme chromatografii, hovoříme o chromatografii afinitní. A právě jednou
z posledních zavedených chromatografických technik je chelatační
afinitní chromatografie, založená na vzájemné interakci mezi biopolymerem v roztoku a kovovými ionty vázanými na pevné fázi,
obvykle hydrofilním polymeru. Porath a kol. publikovali v polovině sedmdesátých let koncept popisující princip
této metody (Porath 1975) a nazvali ji „Metal chelate chromatography“. Název byl
později pozměněn (Porath 1983) na „Immobilized metal ion affinity chromatography“ (IMAC). Vytvoření českého ekvivalentu názvu
metody ponechávám na čtenáři; v tomto textu je používán název Chelatační afinitní chromatografie popř. zkratka IMAC.
Chelatační
sorbent
Historie chelatujících sorbentů je poměrně krátká. První práce v tomto směru pocházejí
ze čtyřicátých let. Od té doby se tato oblast polymerní chemie velice rychle rozvíjí
(viz. přehledné práce Blasius
1963, Schmucker 1977,
Mjaslojedova
1971, Kahovec 1981). Chelatující
sorbent lze v podstatě připravit třemi způsoby.
1/ polykondenzací - chelatační
ligand je obsažen již v některém z monomerů nebo vzniká
až polykondenzací;
2/ polymerací vinylového monomeru
obsahujícího chelatační ligand
3/ zavedením chelatačního ligandu do polymerního nosiče
Polykondenzační postup je už v současné době překonán (produkty
jsou často málo stálé, lze obtížně regulovat stupeň zesítění
a za podmínek polykondenzace může být často napaden samotný chelatační
ligand).
Polymerační způsob naráží na problém přípravy a čištění speciálních
monomerů.
Poslední, třetí způsob
se zdá být nejvhodnější. I tato nejpoužívanější metoda má však své nevýhody. Při
imobilizaci ligandu může totiž docházet k vedlejším reakcím
a mohou vznikat chemicky rozdílná chelatující seskupení.
Nejpoužívanějším polymerním nosičem pro navázání chelatačního ligandu je stejně jako u většiny ostatních afinitních chromatografických technik agarosa. Jsou však využívány i další typy polymerních nosičů jako je sepharosa, supherosa, trisakrylové gely (Boschetti 1983, Moroux 1985) atd. Za zmínku stojí i sorbenty pro vysokotlakou chelatační chromatografii (HP-IMAC), kde je kladen důraz na vysokou mechanickou stabilitu. K tomu účelu byly připraveny nosiče na bázi silikátu (Fanou-Ayi 1983, El-Rassi 1986, Figueroa 1986) popř. na bázi hydrofilních pryskyřic (Kato 1986, Nakagawa 1988, Yip 1989, Belew 1987).
Chelatační
ligand
Nejpoužívanějším ligandem pro imobilizaci na polymerní nosič je iminodiacetát (IDA). V poslední době je však tendence nahrazení IDA ligandem vytvářejícím stabilnější komplexy s kovy. Na sorbentech pro HP-IMAC bylo totiž pozorováno postupné uvolňování kovových iontů ze sorbentu, což má v mnoha případech negativní vliv na separované proteiny. Mezi chelatační ligandy vytvářející stabilnější komplexy s kovy můžeme zařadit karboxymetylaspartát (CM-ASP), tris(karboxymetyl) etylendiamín (TED), N-(2-pyridylmetyl)glycin. Všechny tyto chelatační ligandy byly vhodným způsobem navázány na nosič a následně testovány vlastnosti takto vzniklého chelatačního sorbentu (Monjon 1987, Porath 1988).
komplex chelatační sorbent-kov
Kovové ionty lze rozdělit do tří kategorií
podle jejich reaktivity vůči nukleofilům na tzv. „tvrdé“,
„středně tvrdé“ a „měkké“ (Pearson 1963,
Pearson 1973,
Ho 1977). „Tvrdé“ kovy patří do skupiny silných Lewisových kyselin neboli skupiny silných akceptorů elektronů.
Toto rozdělení nepochybně usnadňuje volbu vhodných kovů využitelných pro IMAC. Hranice mezi jednotlivými kategoriemi jsou však dost neostré
a to hlavně u kovů zařaditelných do skupiny středně silných a slabých Lewisových kyselin, což vede k tomu, že bývají rozlišovány pouze
dvě kategorie kovů: „tvrdé“ a „měkké“. Do kategorie „tvrdých kovů“ lze zařadit ionty
Fe3+, Sc3+, Th4+, Al3+, Ga3+,
In3+. Tyto ionty preferují interakci s atomem kyslíku ligandu. Do kategorie „měkkých kovů“ bývají zařazovány ionty
Cu2+, Zn2+, Ni2+, Co2+, Fe2+,
Mn2+, Mg2+.
Irving a Williams porovnali konstanty stability komplexů těchto kovů
s ligandy obsahujícími kyslík nebo dusík jako donory elektronů
(Irving 1948):
Zn2+ < Cu2+
> Ni2+ > Co2+ > Fe2+ > Mn2+
> Mg2+
Právě ionty „měkkých“
kovů (hlavně Cu2+, Zn2+ a Ni2+) bývají velice často
používány při IMAC.
Při výběru vhodného ligandu a kovu se snažíme, aby komplex ligand-kov
byl co nejstabilnější, na druhé straně však ne tak stabilní, abychom nebyli schopni
za určitých podmínek kov z komplexu uvolnit. Navíc se dá formulovat obecné pravidlo,
Že čím silnější je interakce mezi kovem a ligandem, tím
slabší je pak interakce mezi komplexem ligand-kov a peptidem
(proteinem).
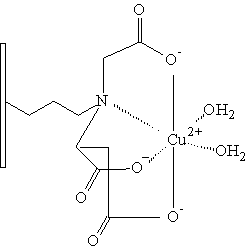
Karboxymethyl aspartát
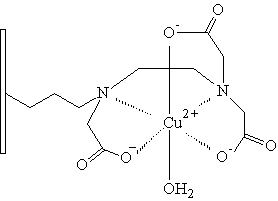
Tris(karboxymethyl) ethylendiamin
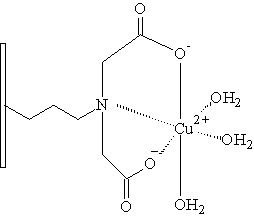
Iminodiacetát
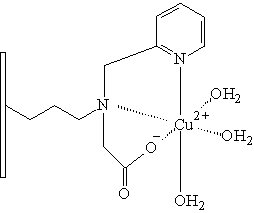
N-(2-pyridylmethyl) glycin
komplex chelatační sorbent-kov-aminokyselina(peptid,protein)
Porath a kol. ve své úvodní
práci (Porath 1975) dali do úzkého vztahu chování proteinů při IMAC a množství imidazolových a thiolových skupin
na povrchu interagujícího proteinu. Také další práce (Porath
1983a, Coulet 1981)
dokazují přítomnost specifické interakce mezi kovem a proteinem, závislé na pH. Tato interakce vynikne po potlačení elektrostatické a hydrofobní
interakce, které také mohou mít vliv na chování proteinu při IMAC. Vliv elektrostatické
interakce se dá odstranit buď použitím vysoké iontové síly v médiu (v převážné většině
prací se používá NaCl v koncentraci od 0.5 do 1 mol/l)
nebo použitím elektroneutrálních komplexů chelatační sorbent-kov. Vliv interakce
hydrofobní je méně zřetelný a prakticky úplně se dá potlačit volbou vhodného chelatačního sorbentu.
Samotná interakce mezi
kovem a proteinem je způsobena vznikem koordinační vazby mezi akceptorem elektronů - kovem a donorem elektronů - aminokyselinovým zbytkem
s vysokou elektronovou hustotou (His, Cys, Trp) orientovaném
na povrchu proteinu (Coulet 1981). Pro tyto ligandy je charakteristická
schopnost stabilizovat nízké oxidační stavy (Cotton
1973).
Na proces interakce je
také možno nahlížet jako na speciální typ Ligand exchange chromatography (LEC), kde
jsou slabě navázané „anti-ligandy“
vody v komplexu chelatační sorbent-kov
nahrazeny „anti-ligandy“ nukleofilnějšími. „Anti-ligandy“ u proteinů a peptidů lze rozdělit do dvou skupin. První
skupinu tvoří aminokyselinové zbytky obsahující imidazol,
thiol nebo indol na povrchu proteinu (peptidu) a druhým
reaktivním centrem je N- terminální aminokyselina
(důležitý je hlavně aminodusík).
Aminokyseliny vytvářejí odlišné komplexy, které mají větší stabilitu než komplexy peptidů nebo proteinů (jejich aminoskupiny jsou silnější base a jejich karboxylové skupiny jsou lepšími donory elektronů než karbonylové skupiny peptidů).
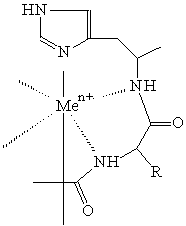
komplex peptidu s His zbytkem
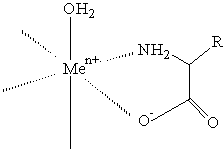
komplex aminokyseliny
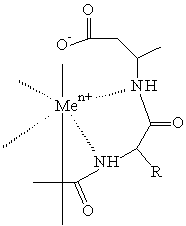
komplex peptidu s COO- zbytkem
Aplikace IMAC
Využití IMAC při purifikaci
Největší využití našla
IMAC, hlavně v počátcích svého vývoje, při purifikaci proteinů krevní plazmy. Tímto
způsobem byl na kolonách s chelatačním sorbentem nasyceným zinkem purifikován makroglobulin
(Sottrup-Jensen
1980, Borth 1981,
Hudson 1982, Imber 1982), inhibitor proteinázy
(Sottrup-Jensen
1980, Kurecki
1979), HS glykoprotein (Lebreton
1977, Lebreton 1979), aktivátory plazminogenu (Rijken 1979,
Rijken 1981). Andersson použil při frakcionaci lidského
séra Sepharosu 6B obsahující ligandy
IDA resp. tris(karboxymetyl)etylendiamín s navázanými ionty Zn2+ a Cd2+.
Lonnerdal a kol. využili IMAC při purifikaci laktoferinu z lidského mléka na chelatačních
sorbentech nasycených mědí. Výtěžek laktoferrinu byl prakticky 100% a navíc biologické vlastnosti
laktoferrinu (inhibice bakteriálního růstu) zůstaly zachovány,
což svědčí o tom, že nedochází k poškození proteinu. Pro dokumentování širokého
spektra použití IMAC lze uvést izolaci interferonu (Knight
1978). Na chelatačních
sorbentech nasycených zinkem, mědí, kobaltem nebo niklem
byly izolovány a charakterizovány interferony z mnoha zdrojů (Egly
1977, Bollin 1978,
Chadha 1979,
Ferreira 1981, Berg 1980,
Berg 1981, Yonehara 1981). Mimo praktické využití při purifikaci dává afinita interferonů
ke kovovým chelátům možnost porovnání jejich strukturálních
podobností, které se odrážejí v orientaci některých aminokyselinových zbytků na
povrchu těchto proteinů (Sulkowski 1982). Smith a kol. (Smith
1987, Smith 1988) využili
s úspěchem faktu, že adsorpce proteinu může být silně zvýrazněna, jestliže jsou
zbytky histidinu lokalizovány blízko N-terminální aminokyseliny peptidického řetězce, k zavedení techniky pro rychlou izolaci
rekombinantních proteinů.
Samostatnou skupinu aplikací
tvoří separace proteinů na chelatačních sorbentech nasycených „tvrdými“ kovy (Fe3+, Al3+,
Ga3+, In3+, Th3+). Jak již bylo uvedeno, tvrdé
kovy preferují při interakci karboxylové a fosfátové skupiny. Byla popsána chromatografie
sérových proteinů (Ramadan 1985a,
Ramadan 1985b), lysozymu, cytochromu c, avidinu, hovězí pankreatické
RNazy, myoglobinu, ovalbuminu (Ramadan 1985c) a specifická sorpce fosfoproteinů
(Andersson 1986). Ve všech případech byl chelatační sorbent nasycen Fe3+.
Další aplikace IMAC
|
Separovaná makromolekula |
Použitý imobilizovaný kovový iont |
odkaz |
|
Nukleosidbisfosfatasa z krysích jater |
Zn2+ |
|
|
Granuloproteiny granulocytů |
Cu2+ |
|
|
Lektin semen Dolichos biflorus |
Zn2+ |
|
|
Gastrointestinální polypeptidy |
Zn2+ |
|
|
Inhibitor trypsinu z lidského séra |
Zn2+ |
|
|
Inhibitor thiol proteinas |
Cu2+ |
|
|
nehistonové proteiny |
Cu2+ |
|
|
nukleotidy |
Cu2+ |
|
|
aminoacylovaná t-RNA |
Cu2+ |
|
|
NADH nitrátreduktasa |
Zn2+ |
|
|
cyt P-450 |
Ni2+ |
|
|
katalasa |
Cu2+ |
|
|
superoxid dismutasa |
Cu2+ |
|
|
Albumin (BSA) |
Cu2+, Zn2+ |
|
|
lysozym |
Cu2+ |
|
|
Ovotransferrin |
Cu2+ |
|
|
Perforin a granzymy lymfocytů |
Co2+ |
|
|
karboxypeptidasa B |
Zn2+ |
Velice důležitou aplikací IMAC se v posledních letech stává jednokroková purifikace rekombinantních proteinů obsahujících navíc takzvanou histidinovou kotvu. Interakce chelatačního nosiče se separovaným proteinem je pak natolik silná, že můžeme podmínky separace nastavit tak, aby v ideálním případě interagoval pouze námi separovaný protein. Imobilizovaným kovovým iontem bývá v tomto případě většinou Ni2+.
Tandemová chromatografie
Chelatační chromatografie se zdá být i velice vhodnou metodou
pro tzv. tandemové uspořádání (při chromatografii je za sebou propojeno několik
kolon naplněných chelatačním sorbentem).
Jednotlivé sorbenty se liší použitým kovem popřípadě na
nosič navázaným ligandem. Je zde tedy velice mnoho kombinací,
které dovolují vytvořit optimální podmínky pro separaci požadovaného proteinu. Tato
idea byla testována například na modelovém vzorku lidských sérových proteinů (Porath
1986,
Porath 1987a,
Porath 1987b). V tomto případě se na první kolonce naplněné chelatačním sorbentem s ligandem TED a nasyceném Zn2+ zachytí při pH 7.0 hlavně makroglobulin a hemopexin, zatímco
převážná většina bílkovin je zachycena na dalších sorbentech
(např Cu-IDA).
Renaturace (refolding)
Jevem, provázejícím expresi proteinů, bývá tvorba nerozpustných útvarů, takzvaných inkluzních tělísek v bakteriální cytoplasmě. Tyto nekorektní konformery rekombinantního proteinu mohou vznikat z mnoha různých důvodů (toxicita rekombinantního proteinu pro bakteriální metabolismus, nepřítomnost vhodného chaperonu atd.). Inkluzní tělíska se dají rozpustit vysokými koncentracemi močoviny nebo guanidinium chloridu a protože IMAC se dá provádět i za denaturačních podmínek, je metoda použitelná i při purifikaci rekombinantního proteinu (s His tagem), který je za nativních podmínek součástí inkluzních tělísek. Zároveň lze vlastní purifikaci využít i pro renaturaci proteinu. V tomto případě je renaturace navázaného proteinu prováděna postupným snižováním koncentrace denaturačního činidla v mobilní fázi. Pokud je tento renaturační protokol dostatečně úspěšný je pak pro nás tvorba inkluzních tělísek spíše výhodou, protože zastoupení rekombinantního proteinu bývá v inkluzních tělíscích velice významná. Navíc jsou inkluzní tělíska snadno oddělitelná od bakteriálního cytosolu.
Příklad renaturačního protokolu a průběhu separace
Využití IMAC při studiu povrchu biomolekul
Jak bylo naznačeno v dřívějším
textu největší vliv na chování biomolekul (proteinů) při
IMAC mají histidinové zbytky orientované na povrchu biomolekuly (v menší míře pak zbytky cysteinu a tryptofanu). V posledních několika letech se dokonce objevují
práce (Muszynska 1986,
Hemdan 1989,
Sulkowski 1985), ve kterých autoři dávají do úzkého vztahu
počet histidinových zbytků na povrchu biomolekuly a sílu interakce mezi biomolekulou
a chelatačním sorbentem. Z výsledků
IMAC se snaží získat informace nejenom o počtu histidinových
zbytků, ale i o „mikrookolí“ histidinu zapojeného do interakce,
popřípadě spojení vlivu více histidinových zbytků orientovaných
dostatečně blízko sebe.
K posouzení těchto interakcí slouží několik základních přístupů:
-
IMAC na chelatačních sorbentech nasycených měďnatými ionty. Chelatační sorbent se ekvilibruje pufrem o pH
-
Chelatační sorbent nasycený měďnatými ionty je ekvilibrován pufrem o pH 7.0 s 1 mmol/l imidazolem a po nanesení vzorku se postupně zvyšuje koncentrace imidazolu v mobilní fázi až na 15 mmol./l. Látky nezachycené (1 mmol/l imidazol) mohou obsahovat jeden histidinový zbytek. Látky s dvěma histidinovými zbytky jsou eluovány 7 - 8 mmol/l imidazolem
-
Chelatační sorbent je nasycen nikelnatými ionty a po nanesení vzorku jsou látky izokraticky eluovány startovním pufrem o pH 7.0. V mrtvém objemu kolony vytékají látky obsahující jeden nebo neobsahující žádný histidinový zbytek. V desetinásobku mrtvého objemu eluují látky se dvěma histidinovými zbytky. V šedesátinásobku mrtvého objemu pak látky se čtyřmi popřípadě pěti histidinovými zbytky
Nabízí se i možnost „odfiltrování“
interagujících aminokyselinových zbytků specifickou
modifikací. Tato metoda by mohla být zvlášť vhodná při studiu vlivu jiných
aminokyselinových zbytků, než je histidin. Vliv těchto aminokyselinových zbytků
je v přítomnosti histidinu malý. Bylo popsáno chování ovotransferrinu
při IMAC po modifikaci histidinových zbytků diethylpyrokarbonátem (Al-Mashikhi
1987). V tomto případě se modifikovaný ovotransferrin na chelatačním sorbentu nasyceném mědí vůbec nezachytil. E. Sulkowski (Sulkowski 1987) sledoval chování chymotrypsinogenu
A, chymotrypsinu A a chymotrypsinu při IMAC na chelatačním
sorbentu nasyceném mědí. Zjistil veliké rozdíly v eluci těchto látek po přidání diizopropylfosfofluoridátu.
Tyto změny vysvětlil změnou „mikrookolí“ interagujícího histidinu.
Příklady interakce kov-protein
1.Apoferritin
Apoferritin je intracelulární protein, který
váže pro buňku toxické
Fe3+ ionty a zároveň slouží jako rezervní zdroj železa pro syntézu cytochromů
a dalších Fe-obsahujících biomolekul.
Molekula apoferritinu je schopna vyvázat až 4500 atomů
železa (Theil 1987,
Chasteen 1985). Zdá se tedy velice pravděpodobné, že alespoň část z vazebných míst
pro železo je umístěna blízko povrchu nebo přímo na povrchu apoferritinu.
Chelatační chromatografie na sorbentech
nasycených Fe3+ tedy může být vhodnou metodou nejenom pro purifikaci
apoferritinu, ale i pro studium aktivního místa. R. F.
Boyer a kol. (Boyer 1990) zjistili, že vazba do tohoto místa je
závislá na pH - nejsilnější interakce byla pozorována při pH 4.0. Tato skutečnost naznačuje, že na interakci s Fe3+
se podílí hlavně ligandy s kyslíkovým atomem. V aktivních
místech byla prokázána přítomnost zbytků kyselin asparagové
a glutamové (Yang
1987, Sayers 1983,
Wardeska 1986). Komplex
apoferritin : Fe3+ je stabilní v přítomnosti
1 mol/l NaCl. V
případě použití silného chelatačního činidla, jako je
oxalát, citrát nebo EDTA je vazba chelatační ligand : Fe3+
přerušena a dochází k eluci komplexu apoferritin : Fe3+. K eluci
apoferritinu bez Fe3+ dochází pouze zvýšením
pH nebo vlivem fosfátu popřípadě fosfoserinu
(Boyer
1990).
Závěry:
A/ Větší afinita apoferritinu k chelatačnímu sorbentu
nasycenému Fe3+
při nižších pH indikuje důležitost iontových interakcí
(Belew 1987, Ramadan 1985c
)
B/ Stabilita komplexu
apoferritin : Fe3+ v 1 mol/l NaCl signalizuje, že mimo iontových interakcí dochází i ke vzniku
koordinačně-kovalentních vazeb (Boyer
1990)
2. Albumin (BSA)
Albumin je velice zajímavá
biomolekula, jejíž funkcí v organismu je mimo jiné i transport
širokého spektra látek (Feihke
1979, Gambhir 1975,
Jacobson 1969,
Wilting 1981, Nakano 1982). Vazba Cu2+ na
BSA byla studována v mnoha laboratořích (Kure
1984, Nakano 1982,
Mohanakrishnan 1982,
Klotz 1948, Naik 1975,
Katz 1980). Při pH 7.4 byly u BSA nalezeny tři místa schopné více či méně silně
vázat Cu2+ (Zgirski
1990). Vazebné místo 1 je lokalizováno na N-terminálním konci
proteinu. Obsahuje tedy aminodusík N-terminální aminokyseliny
(Asp), imidazolový dusík histidinového zbytku v pozici 3, dva dusíky peptidické vazby (Thr{2}, His{3})
a kyslík karboxylu (Asp) (Schearer
1967, Sarkar 1982,
Laussac 1984).
Asociační konstanta (Ka) pro komplex Cu2+ :
vazebné místo 1 se pohybuje v rozsahu 1.1 x 1012 -
1.6 x 1013 mol-1 (Giroux
1981, Syvertsen 1986). Cu2+ není odstranitelný
chelatačně ani při velmi nízkých průtocích (3 ml/hod)
na chelatačních sorbentech nenasycených
kovem. Vazebné místo 2. Struktura vazebného místa 2 nebyla dosud dostatečně prostudována.
Cu2+ se do tohoto místa váže slaběji a je neodstranitelný chelatačně pouze při vysokých průtocích (40 ml/hod). Asociační
konstanta se v tomto případě pohybuje kolem 5.2 x 106 mol-1
(Zgirski
1990, Schearer 1967,
Syvertsen 1986). Při pH 7.4 je BSA schopen vyvázat další tři
ionty Cu2+. Asociační konstanta je v tomto případě ještě nižší a pohybuje
se kolem hodnoty 1.6 x 105 mol-1. Tyto atomy mědi jsou sice
neoddialyzovatelné, ale odstranitelné chelatačně i při velmi vysokých průtocích (Zgirski
1990). Je však nutné
zdůraznit, že množství vazebných míst pro Cu2+ na BSA je značně závislé
na pH. Při pH 4.8 - 5.3 byla nalezena 1 - 3 místa s Ka
0.8 - 30 x 105 mol-
Literatura
1.
J. Porath,
J. Carlsson, I. Olin, G. Belfrage,
Nature (London), 258, 598 (1975).
2.
J. Porath, I. Olin,
Biochemistry, 22, 1621 (1983).
3.
E. Blasius, B. Brozio, H. A. Flaschka, A. J. Barnard (eds.), Chelates in Analytical Chemistry, 1. díl, M. Dekker, New York 1963.
4.
G. Schmucker, v knize:
Encyklopedia of Polymer Science
and Technology, Supplement,
2. díl, str.197. Interscience,
New York, London, Sydney, Toronto 1977.
5.
G. V. Mjaslojedova,
O. P. Jenisejeva, S. B. Savin,
Žur. Anal. Chim., 26, 2172 (1971).
6.
J. Kahovec, Chem. listy, 75, 398 (1981).
7.
E. Boschetti, P. Girot, O. Pepin, Sci. Tools, 30, 27 (1983).
8.
Y. Moroux, E. Boschetti, J. M. Egly, Sci. Tools, 32, 1 (1985).
9.
L. Fanou-Ayi, M. Vijalakshmi, Ann. NY. Acad. Sci, 413, 300 (1983).
10.
Z. El-Rassi, C. Horvath, J. Chromatog., 359, 241 (1986).
11.
A. Figueroa, C. Corradini, B. Feibush, B. Karger, J. Chromatogr., 371, 335 (1986).
12.
Y. Kato, K. Nakamura,
T. Hashimoto, J. Chromatogr.,
354 511 (1986).
13.
Y. Nakagawa, T. T. Yip, M. Belew, J. Porath, Anal. Biochem.,
168, 75 (1988).
14.
T. T. Yip, Y. Nakagawa, J. Porath, Anal. Biochem., 183 159 (1989).
15.
M. Belew, T. T. Yip, L. Andersson, R. Ehrstrom, Anal. Biochem., 164, 457 (1987).
16.
B. Monjon, J. Solms, Anal. Biochem.,
160, 88 (1987).
17.
J. Porath, Trends in Anal. Chem., 7, 254 (1988).
18.
R. G. Pearson, J. Am. Chem. Soc., 85, 3533 (1963).
19.
R. G. Pearson, Hard and soft acids
and bases Dowden, Hutchinson, Ross, Stroudsberg, 1973.
20.
T. L. Ho, Hard and soft acids
and bases, Principles in Organic chemistry, Acadedic Press, New York, 1977.
21.
H. Irving, R.J.P. Williams, Nature (London), 162, 746 (1948).
22.
J. Porath, M. Belew, v knize: Affinity chromatography and biological recognition, I. M. Chaiken, M. Wilchek, I. Parikh, Eds., str. 173. Academic Press, New York, 1983.
23.
P. R. Coulet, J. Carlsson, J. Porath, Biotechnol. Bioeng. 23, 663 (1981).
24.
F. A. Cotton, G. Wilkinson, v knize: Anorganická
chemie, Academia, str. 691. Praha, 1973.
25.
L. Sottrup-Jensen, T. E. Petersen, S. Magnusson, FEBS Lett., 121, 275 (1980).
26.
W. Borth, E. J. Menzel, M. Salzer, C. Steffen, Clin. Chim. Acta, 117, 219 (1981).
27.
N. W. Hudson, P. H.
Koo, Biochim. Biophys. Acta, 704 290 (1982).
28.
M. J. Imber, S. V. Pizzo,
W. J. Johnson, D. O. Adams,
J. Biol. Chem., 257, 5129 (1982).
29.
T. Kurecki, L. E. Kress, M. Laskowski, Anal. Biochem. 99, 415 (1979).
30.
J. P. Lebreton, FEBS
Lett., 80, 351 (1977).
31.
J. P. Lebreton, F. Joisel, J. P. Rault, J. Clin. Invest. 64, 1118 (1979).
32.
D. C. Rijken, G. Wijngaards, M. Z. DeJong, J. Welbergen Biochim. Biophys. Acta, 580, 140 (1979).
33.
D. C. Rijken, D. Collen, J. Biol. Chem., 256, 7035 (1981).
34.
L. Andersson, J. Chromatogr., 315, 167 (1984).
35.
B. Lonnerdal, J. Carlsson, J. Porath, FEBS Lett., 75, 89 (1977).
36.
E. Knight, J. Pharmac. Ther. A., 2, 439 (1978).
37.
V. G. Egly, A. Billiau, P. DeSomer, J. Biol. Chem., 252 5934 (1977).
38.
E. Bollin, E. Sulkovski, Arch. Virol., 58, 149 (1978).
39.
K. C. Chadha, P. M.
Grob, A. J. Mikulski, L. R.
Davis, E. Sulkowski, J. Gen.
Virol., 43, 701 (1979).
40.
P. C. P. Ferreira, M.
Paucher, R. R. Golgher, Arch.
Virol., 68, 27 (1981).
41.
K. Berg, I. Heron, J. Gen. Virol.(London), 50,
441 (1981).
42.
K. Berg, I. Heron, Scand. J. Immunol., 11, 489 (1980).
43.
S. Yonehara, Y. Yanase, T. Sano, M. Imai, S. Nakasawa, H. Mori, J. Biol. Chem., 256, 3770 (1981).
44.
E. Sulkowski, K. Vastola a D. Oleszek, v knize: Affinity Chromatography and Related Techniqes,
T. C. J. Gribnau, J. Visser,
R. J. F. Nivard, Eds., str.
313. Veldhoven, The Netherlands, 1982.
45.
M. C. Smith, T. C. Furman,
C. Pidgeon, Inorg. Chem., 26 1965 (1987).
46.
M. C. Smith, T. C. Furman,
T. D. Ingolia, C. Pidgeon, J.
Biol. Chem., 263, 7211 (1988).
47.
N. Ramadan, J. Porath,
J. Chromatogr., 321, 81 (1985).
48.
N. Ramadan, J. Porath,
J. Chromatogr., 321, 105 (1985).
49.
N. Ramadan, J. Porath,
J. Chromatogr., 321, 93 (1985).
50.
L. Andersson, J. Porath, Anal. Biochem.,
154, 250 (1986).
51.
I. Ohkubo, T. Kondo, N. Taniguchi, Biochim. Biophys. Acta, 616, 89 (1980).
52.
A. R. Tores, E. A. Peterson, W. H. Evans, M. G. Mage, S. M. Wilson, Biochim. Biophys. Acta, 576, 385 (1979).
53.
C. A. K. Borrebaeck,
B. Lonnerdal, M. E. Etzler,
FEBS Lett., 130, 194 (1981).
54.
M. Conlon, R. F. Murphy, Biochem. Soc. Trans., 4, 865
(1976).
55.
J. P. Salier, J. P.
56.
H. C. Ryley, Biochem. Biophys. Res. Commun., 89, 871 (1979).
57.
H. Kikuchi, M. Watanabe, Anal. Biochem., 115, 109 (1981).
58.
P. Hubert, J. Porath,
J. Chromatogr., 206 (1981) 164
59.
A. J. Gozdzicka-Jozefiak, D. Augustyniak, J. Chromatogr., 131, 91 (1977).
60.
M. G. Redinbaugh, W.
H. Campbell, Plant Physiol., 71, 205 (1983).
61.
M. Kastner, D. Neubert, J. Chromatogr., 587, 43 (1991)
62.
M. Miyata-Asano, K. Ito, H. Ikeda, S. Sekiguchi, K. Arai, N. Taniguchi, J. Chromatogr., 370, 501 (1986).
63.
R. J. Weselake, S. L.
Chesney, A. Petkau, A. D. Friesen, Anal. Biochem., 155, 193 (1986).
64. H. Hansson, L. Kagedal, Adsorption and desorption of proteins in metal chelate affinity chromatography : Purification of albumin. J. Chromatogr., 215, 333 - 339 (1981).
65.
A. R. Torres, E. A.
Peterson, W. H. Evans, M. G.
Mage, S. M. Wilson, Biochim. Biophys. Acta, 576, 385 (1979)
66.
S. Al-Mashikhi, S. Nakai, Agric Biol. Chem.,
51, 2881 (1987).
67.
U. Winkler, T.M. Pickett, D. Hudig, Fractionation of perforin and granzymes
by immobilized metal affinity
chromatography, Journal of Imunological Methods, 191, 11 (1996).
68.
G. L. Hortin, B. L. Gibson, Purification
of carboxypeptidase B by zinc chelate
chromatography, Prep. Biochem., 19, 49 (1989).
69.
G. Muszynska, L Andersson, J. Porath, Biochemistry, 25 6856 (1986).
70.
E. S. Hemdan, Y. Zhao, E. Sulkowski, J. Porath, Proc. Natl.
Acad. Sci. USA, 86, 1811 (1989).
71.
E. Sulkowski, Trends Biotechnol., 3, 1 (1985).
72.
E. Sulkowski, v knize:
Burgess R. ed. Protein purification: micro to macro, Burgess R. ed., str. 149. New York, A. R. Liss, 1987.
73.
E. C. Theil, Annu. Rev. Biochem., 56, 289 (1987).
74.
N. D. Chasteen, B. C.
Antanaitis, P. Aisen, J. Biol. Chem.,260, 2926 (1985).
75.
R. F. Boyer, S. M. Generous, T. J. Nieuwenhuis, R. A.
Ettinger, Biotechnol. Appl. Biochem., 12, 79 (1990).
76.
C. Yang, A. Meagher, B. H. Huynh, D. E. Sayers, E. C. Theil, Biochemistry, 26, 497 (1987).
77.
D. E. Sayers, E. C.
Theil, F. J. Rennick, J. Biol. Chem. 258, 14076 (1983).
78.
J. G. Wardeska, B. Viglione, N. D. Chasteen, J. Biol. Chem., 261, 6677 (1986).
79.
N. D. Chasteen, E. C.
Theil, J. Biol. Chem., 257, 7672 (1982).
80.
A. Treffry, P. M. Harrison,
J. Inorg. Biochem., 21, 9 (1984).
81.
K. J. Feihke, W. E.
Muller, U. Wollert, Biochim. Biophys. Acta, 557, 346 (1979).
82.
K. K. Gambhir, R. H.
Menamy, F. Watson, J. Biol. Chem. 250, 6711 (1975).
83.
J. Jacobson, FEBS Lett., 5, 112 (1969).
84.
J. Wilting, W. F. Van
der Giesen, L. H. M. Janssen,
Bioch. Pharmacol., 10, 1025
(1981).
85.
N. I. Nakano, Y. Shimamori, S. Yamaguchi, J. Chromatogr. 237, 225 (1982).
86.
P. Mohanakrishnan, C.
F. Chignell, J. Pharm. Sci., 71 1180 (1982).
87.
N. Kure, H. Nakatsuji,
T. Sano, T. Yasunaga, Bull.
Chem. Soc. Jpn., 57, 2712 (1984).
88.
I. M. Klotz, H. G. Curme, J. Am. Chem.
Soc., 70, 939 (1948).
89.
D. V. Naik, C. F. Jewell, S. G. Schulman, J. Pharm. Sci., 64, 1243 (1975).
90.
S. Katz, R. G. Shinaberry, E. Heck, W. Squire, Biochemistry, 19, 3805 (1980).
91.
A. Zgirski, E. Frieden, J. Inorg. Biochem., 39, 137 (1990).
92.
W. T. Schearer, R. A.
Bradshaw, F. R. N. Gurd, J.
Biol. Chem., 242, 5451 (1967).
93.
B. Sarkar, J. Indian Chem. Soc., 59, 1403 (1982).
94.
J. P. Laussac, B. Sarkar, Characterization of the copper(II)-
and nickel(II)-transport
site of human
serum albumin. Studies of copper(II)- and nickel(II) binding to peptide 1-24 of human serum albumin by 13C
and 1H NMR spectroscopy.
Biochemistry, 23, 2832-2838 (1984).
95.
E. Giroux, J. Schoun, J. Inorg. Biochem., 14, 359 (1981).
96.
Ch. Syvertsen, R. Gaustad,
K. Schoeder, T. Ljones, J. Inorg. Biochem., 26, 63 (1986).
97.
I. M. Kolthoff, B. R.
Willeford, J. Am. Chem. Soc., 79 2656 (1957).
98.
J. Porath, J. Chromatogr., 376, 331 (1986).
99.
J. Porath, Biopolymers, 26, 193 (1987).
100. J. Porath, Biotechnol. Progr., 3, 14 (1987).